
Introduction
Since the first probes confirmed life in Proxima b over 100 years ago, many others were sent to study the planet’s diverse life forms and their biology. All information contained in this page has been gathered via probe-based missions. There are a lot of assumptions particularly around evolution, made entirely on observation of living creatures without analyzing their fossil record (if there is one). All of these will be validated and updated after Noah-1 arrives.
Autotrophs
Similarly to life on Earth, Pb’s bottom of the food chain is composed of autotroph organisms, i.e., who can synthesize their own food. There are three major branches:
Photosynthetic organisms, called photo-autotrophs, use light to power their metabolism. These are analogous to plants and algae on Earth, but most species use a pigment called phaeophytin to capture light, which makes them look purple. Photo-autotrophs exist in daylight areas only (Day Hemisphere and Terminator Line) since they need light to survive. Exposure to light per time unit is comparatively less than Earth plants since Proxima Centauri’s light is fainter than the Sun’s. However, these organisms are in a permanent day due to tidal locking, which increases their energy and food production.
Chemosynthetic organisms, called chemo-autotrophs, perform chemical reactions in the absence of light to produce their food. They exist in the Night Hemisphere. Chemical reactions are less energy-efficient than photosynthesis, which is the reason why analogous chemosynthetic organisms on Earth are limited to bacteria. On Proxima b however, chemo-autotrophs do not compete with photo-autotrophs since the latter cannot survive in the Night Hemisphere. In the absence of more energy-efficient competition, chemo-autotrophs evolved into more complex life forms that dominate the bottom of the food chain in the Night Hemisphere (oceans and land).
Radio-synthetic organisms, or radio-autotrophs, use radiation to power their metabolism. It’s a highly efficient but unstable way to generate energy. Only a few plant-like organisms are known in the South Pole, at the edge of the Dead Zone, before radiation becomes too strong for life to exist.
Heterotrophs
Unlike autotrophs who can synthesize their own food, heterotrophs need to eat their food. They are either herbivore (if they eat autotrophs) or carnivore (if they eat other heterotrophs). They are analogous to animals on Earth life, so the terms “heterotroph” and “animal” mean the same thing on our articles.
Proxima b’s heterotrophs evolved into many different groups with different characteristics depending on the hemisphere where they originated. Species native to daylight habitats use light-based vision to perceive their surroundings, while species native to the Night Hemisphere use a combination of sonar-based echolocation and thermal vision. Most animals across hemispheres can hear and smell.






Animals at the top of the food chain are:
Endocrustae are a diverse branch of animals with a plated endoskeleton, evolved from what was previously an exoskeleton in early invertebrate ancestors. We believe these to be the earliest animal group to have invaded the land since they currently dominate land habitats. Mantises and Grabbers are examples of Endocrustae animals.
Bicephalia, predators with two differentiated heads. The sensing head contains sensory organs (eyes, ears, smell sensors) while the feeding head includes the mouth and organs used to capture and kill prey. Squid and Snappers are the common examples. They seem to have appeared in the oceans first and invaded land after Endocrustae were already well established.
Sonosentios, animals native to the Night Hemisphere that detect their surroundings using echolocation and thermal vision. They have a sonar instead of eyes.
Biochemistry
All multicellular organisms in Proxima b have carbon-based biochemistry and use sulfuric acid as a solvent since this is the most abundant liquid on the planet. Basic building blocks are protein-like carbohydrate molecules, i.e., composed of carbon, hydrogen, and oxygen. Many of these molecules also contain sulfur and nitrogen, or tungsten and silica for harder structures such as bones and shells.
Proxima b’s life proteins are incompatible with Earth life proteins. They interact with sulfuric acid instead of water. As such they react very strongly with water - pouring water over a Proxima b life form would have the same effect as pouring sulfuric acid over an Earth life form. Because of this incompatibility, Noah-1 settlers will not be able to consume Proxima b life forms and will instead need to synthesize their own food.
Biological activity creates a carbon and sulfur cycle, similar to Earth’s carbon cycle, where carbon and sulfur are used and restored continuously.
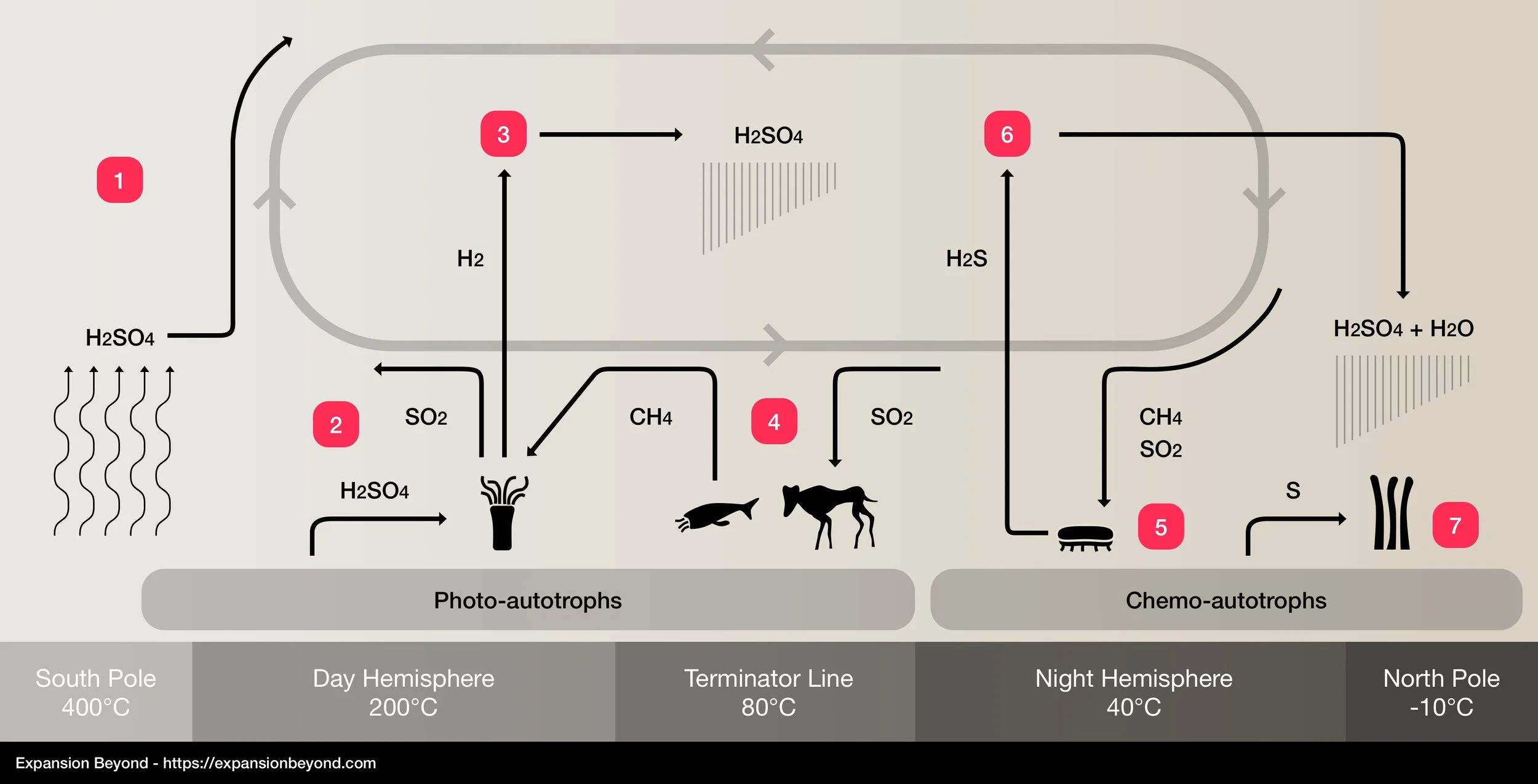
Proxima b’s carbon and sulfur cycle
The carbon and sulfur cycle works as follows:
1. Proxima b’s tidal locking causes the atmosphere to continually flow from the South to the North poles due to temperature differences. In the hottest area of the South Pole (called the Dead Zone), temperatures rise over sulfuric acid’s boiling point, causing it to evaporate into the atmosphere.
2. Photosynthetic organisms use light emitted by Proxima Centauri to power chemical reactions between methane (extracted from the atmosphere) and sulfuric acid (obtained directly from the ocean, rivers, rains or underground veins). This reaction produces a carbohydrate called glycerol, which is later used to produce energy and synthesize other structural carbon-based molecules. This reaction also produces sulfur dioxide and hydrogen, both of which are released back into the atmosphere.
6CH4 + 3H2SO4 = 2C3H8O3 + 3SO2 + 15H2
3. Hydrogen released by photosynthesis reacts with sulfur dioxide in the atmosphere, forming sulfuric acid which condenses and falls as rain. This reaction is the most significant contributor towards rain in the Day Hemisphere.
4. Animals breathe sulfur dioxide and release methane, reversing the effects of photosynthesis. Being heterotrophs, they do not synthesize carbohydrate compounds and instead obtain them by eating autotrophs (if herbivore) or other heterotrophs (if carnivore).
5. Chemosynthetic organisms consume methane and sulfur dioxide from the atmosphere to produce the carbohydrate erythrose, with hydrogen sulfide and hydrogen also being released as part of this reaction. There is enough energy in sulfur dioxide to react with methane without light, which is why chemo-autotrophs can survive in the dark. Their dependence on sulfur dioxide indicates that photo-autotrophs must have appeared first, creating the atmospheric conditions necessary for chemo-autotrophs to function.
4CH4 + 2SO2 = C4H8O4 + 2H2S + 2H2
6. Hydrogen sulfide released into the atmosphere as part of chemosynthesis reacts with sulfur dioxide to form water and sulfuric acid, both of which mix and condense into acid rains. The Night Hemisphere is almost always under acid rains.
7. Advanced, land-based chemo-autotrophs capture elemental sulfur and mix it with methane and rainwater to form methanol and hydrogen sulfide.
S + CH4 + H2O = CH3OH + H2S
Although the macro-biochemistry described above is understood, more investigation is required to learn about metabolic reactions that power cellular respiration. We know that ATP (adenosine triphosphate), the universal energy-carrying molecule in Earth life, is not used by Proxima b’s life forms since it has not been detected in any biological sample taken to date. Genetic information carriers, analogous to Earth life DNA, are also unknown. Genes are clearly carried and passed on to other generations via reproduction, however, cells of Proxima b’s multicellular organisms don’t have a nucleus or other obvious places where genetic information would be carried. At SEV’s Biological Labs, we are looking forward to Noah-1’s arrival, which will provide us with biological samples and live specimens necessary to unlock these secrets.
Feeding and digestion
Proxima b animals need to eat food just like Earth animals. Their primary digestive system structure is similar, consisting of a tube where food is broken down so that nutrients can be absorbed and carried by blood cells, after which the remaining waste materials are expelled. Proxima b’s animal species diverged early into two very distinct branches:
Praeostia: prae (in front) and ostium (mouth), is a branch where the mouth is located at the front of the body, along with sensing organs (vision, hearing, and smell) and organs used to kill prey in the case of predatory species. Eating, digesting and defecation happen in the same direction that the animal move. This is the same anatomy that Earth animals have. Examples of Praeostia species are Squid and Snappers.
Tergumostia: tergum (back) and ostium (mouth), is a branch where the mouth is located at the back of the body, while senses and feeding organs are located in the front of the body. This means that eating, digesting and excreting happen in the opposite direction in which the animal moves. There are no animals with this anatomy on Earth. All Endocrustae species such as Mantises belong to this branch, along with smaller organisms with an exoskeleton.
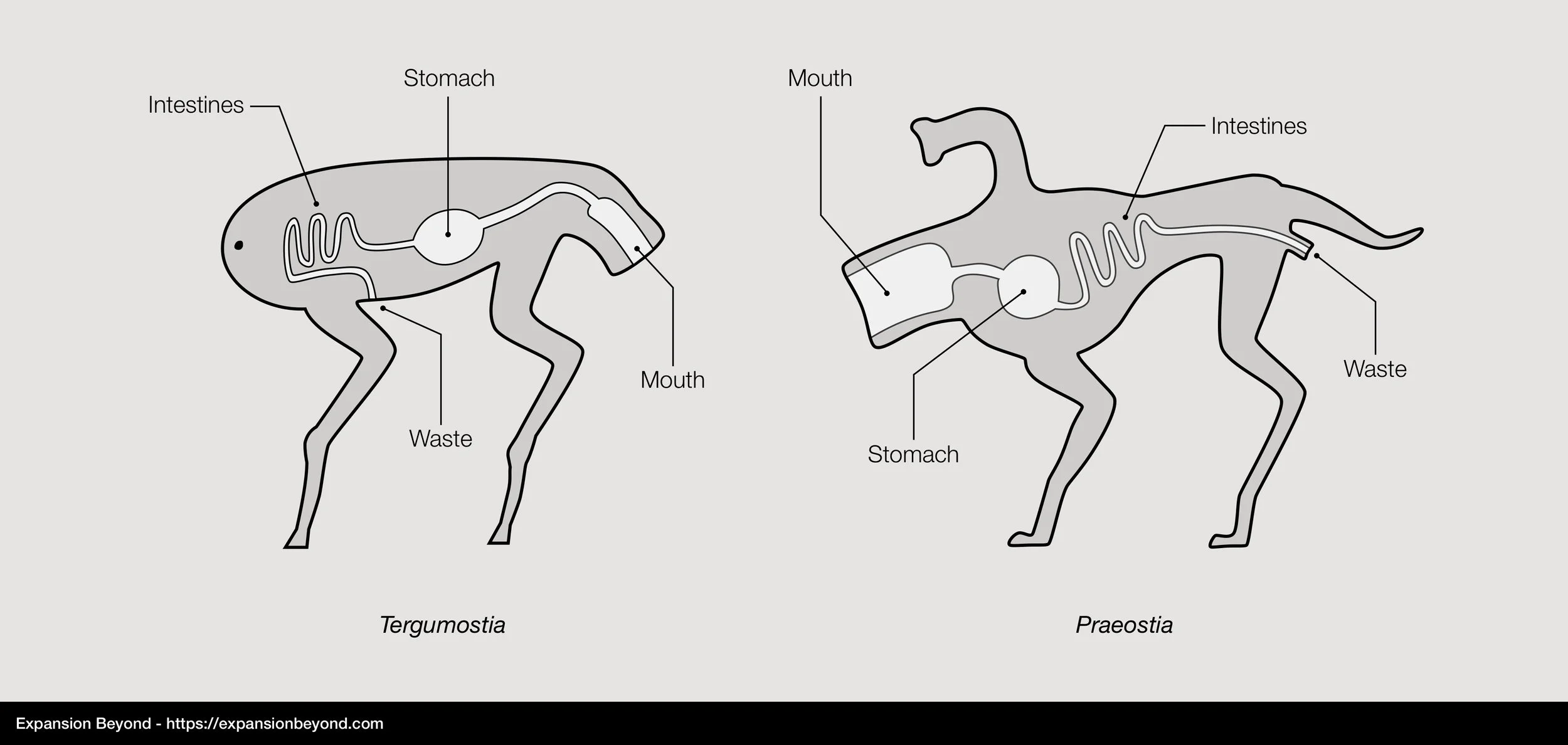
Differences in Tergumostia and Praeostia digestive systems
The Tergumostia branch is older, having appeared in the oceans before Praeostia, and subsequently expanded to land before Praeostia species started to emerge. We believe the mouth at the back of the body is an adaptation to bottom feeding behavior in liquid environments, which initially gave this branch an evolutionary advantage over Praeostia. See taxonomy to learn about how each of these branches evolved separately.
Respiration and circulation
Proxima b animals need to breathe sulfur dioxide (SO2) to perform their metabolic functions, but in much lesser quantities than Earth animals need to breathe oxygen. Reasons for this are not yet understood since we need to learn more about their cellular respiration. But we do know that lower respiratory needs shaped their respiratory and circulatory systems very differently from Earth animals.
To start with, Proxima b animals have an open circulatory system. Earth vertebrates have a closed circulatory system, that is, blood is kept separate from surrounding tissue by flowing through arteries and veins, pumped by the heart. Proxima b animals don’t have arteries or veins; instead, blood is free to flow between body cavities. They do have organs with the same function as a heart (multiple in fact), forcing the blood to circulate across the entire body. Blood, composed of specialized carrying cells diluted in sulfuric acid, is used to exchange gases (sulfur dioxide in, methane out), deliver nutrients to surrounding tissues, and carry wasteful byproducts of cellular respiration into excretion organs. The open circulatory system is much less efficient at delivering sulfur dioxide than Earth vertebrates’ closed circulatory system at delivering oxygen. However, due to Proxima b’s animals’ comparatively lower need for sulfur dioxide and higher concentrations of that gas in the atmosphere, the open circulatory system is sufficient for their survival. Despite its lower efficiency, the open circulatory system makes Proxima b animals much less likely to die by bleeding out. In the event of injury, blood loss is slower since it is not pushed out by the arterial pressure that exists in closed circulatory systems.
Animals’ respiratory system is also adapted for lower gas exchange needs, consisting of a simple tubular network more comparable to Earth insects’ tracheas than vertebrate lungs (although the trachea network is less dense in comparison with Earth insects). For this reason, their respiratory tubes are also called tracheas. Air is inhaled and stays inside the trachea network for enough time for gas exchanges to happen between tiny ramifications of the trachea and surrounding blood. Once the exchange happens, the air is exhaled, and the cycle repeats. Mid-size animals breathe an average of twice per minute, which is 5 to 10 times slower than similarly sized Earth animals.
All of the above is true for both Tergumostia (mouth at the back) and Praeostia (mouth at the front) branches, but there are a few differences in their anatomy:
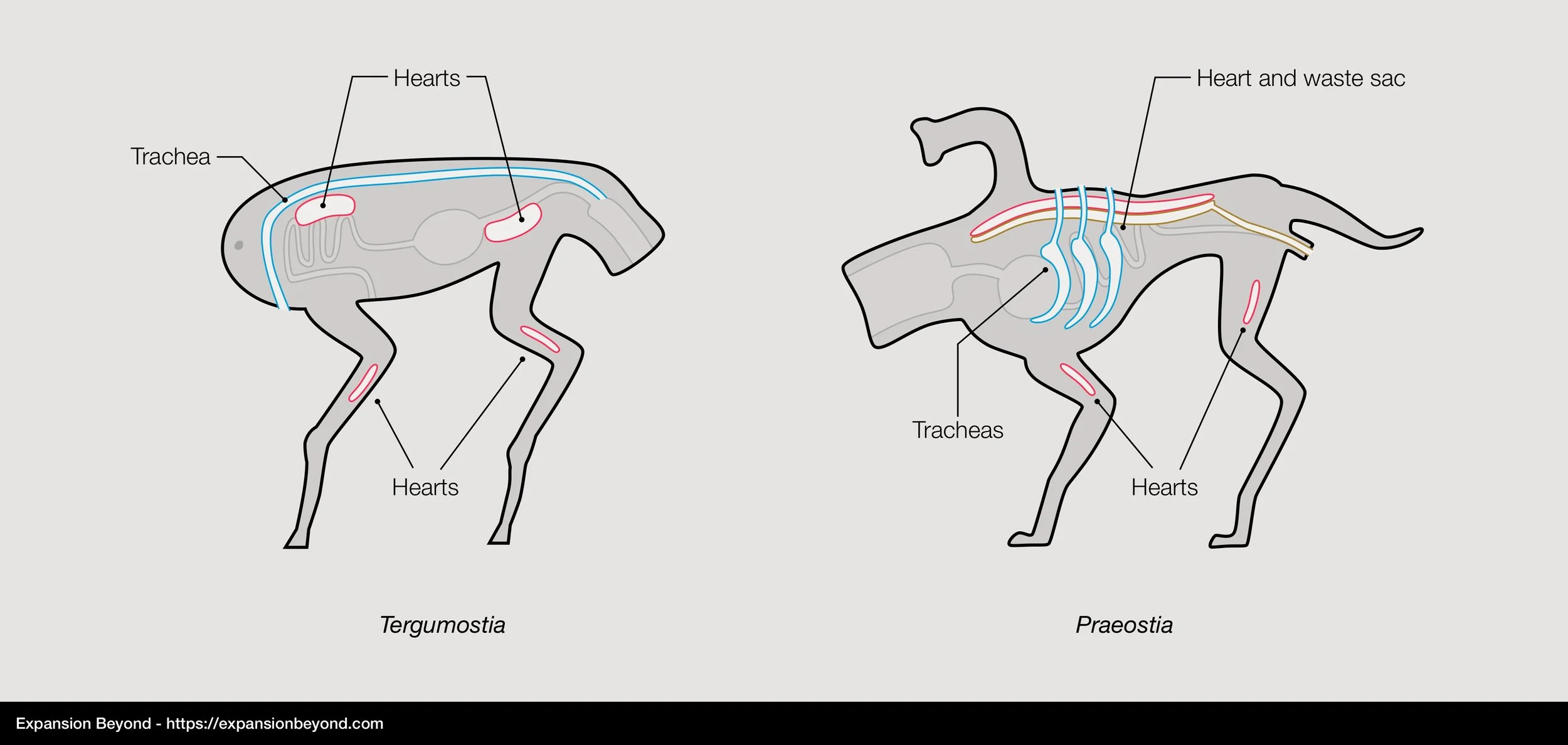
Differences in Tergumostia and Praeostia respiratory and circulatory systems
Tracheas: Tergumostia have a single trachea that goes across their bodies, where air is inhaled through an opening in the front (called the spire) and exhaled through the mouth. Praeostia have multiple spires (organized in either three or four pairs, usually on their backs) from which air is both inhaled and exhaled.
Hearts: Tergumostia have two main hearts on each end of their bodies, plus one smaller heart in each leg that ensures blood reaches their extremities. Praeostia have one main, elongated heart located along their spine which keeps blood circulating in a circular motion from back to front. They also have one small heart in each leg, similar to Tergumostia. We believe this similarity is due to convergent evolution rather than inheritance from a common ancestor.
Waste extraction: Tergumostia don’t have kidneys. Waste products resulting from cellular respiration are collected by the outer walls of their intestines and excreted along with residues from digestion. On Praeostia species, the main heart also performs the function of a kidney. As blood flows through the main heart, it filters waste products and collects it in the waste sac (shown in the diagram above in a different color, but it’s actually the same organ). Waste products are then channeled to the lower intestine where it joins results of digestion to be excreted.
Reproduction
Tergumostia and Praeostia also differ significantly in how they reproduce. Tergumostia have only one gender, and for most species mating means exchanging reproductive cells such that each individual can fertilize each other. Most Tergumostia species are viviparous, and the most significant divergence occurred between Scalprutora (scorpions) and other branches. Scalprutora carry their eggs and give birth through small pouches located in their backs, while all other branches have internal reproductive organs and give birth through their mouth.
Praeostia have two genders, although their definition is different from Earth animals. On Earth, males produce a reproductive fluid (the semen), and females produce eggs. Males fertilize the female's eggs, while the female either lays eggs externally or carries them internally (by developing an embryo) and later gives birth. On Praeostia species, one gender also produces semen while the other produces eggs; however, the gender that produces semen is the one who lays eggs or gives birth. To differentiate the two genders, we use the following definition:
- Males are the gender which produces eggs.
- Females are the gender which produces semen, receives the male's eggs and then lays fertilized eggs or give birth. Details of each species' reproductive strategy are documented in the Life Catalog section.
Content in this page is maintained by our Biological Research Division. We will keep posting updates as we learn more about Proxima b life forms.